Zellzyklus
 Zellteilung verteilt identische Chromosomen auf die Tochterzellen.
Zellteilung verteilt identische Chromosomen auf die Tochterzellen.
Genom = Ausstattung der Zelle mit DNA, der genetischen Information
Somazellen = alle Kperzellen ausser den Gameten, diploid
Gameten = Geschlechtszellen (Eizellen & Spermien), haploid
Chromatin = DNA-Protein-Komplex, kontrolliert die Aktivit舩 der Gene
Mitose = Kernteilung
Cytokinese = Teilung des Cytoplasma
Meiose = Variation der Mitose, bei der die Tochterzellen nur die Häfte der elterlichen Chromosomen erhalten; geschieht nur in den Geschlechtsorganen
Der mitotische Zellzyklus
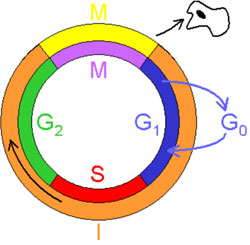
Im Zellzyklus wechseln Mitose und Interphase ab: Eine Übersicht
Die Interphase dauert die meiste Zeit des Zellzyklus, sie wird in unterschiedliche Phasen unterteilt:
- G1-Phase: die Zelle wächst, bereitet die nachfolgende Chromosomenverdoppelung vor
- S-Phase: Chromosomen werden verdoppelt (Synthese von DNA)
- G2-Phase: die Zelle wächst weiter, bereitet die nächste Mitose vor
Die mitotische Phase (M-Phase) beinhaltet die Mitose und die Cytoskinese. Die Mitose selber ist auch wieder in verschiedene Phasen aufgeteilt:
- Prophase: Die Chromosomen werden als lange, düne
Fäden sichtbar da sie sich zur Transportform verkürzen;
Centriol und Centrosomen trennen sich und wandern zu entgegengesetzten Polen; die Kernspindel formt sich und der Nucleolus verschwindet
- Prometaphase: Die Kernmembran wird zerstückelt; Bündel
von Mikrotubuli gehen von den Polen in Richtung Mitte, wo sie sich an den Kinetochoren (die in der Centromerregion der Chromosomen angelagert sind) anlagern; Mikrotubuli, die nicht mit Kinetochoren
Kontakt aufnehmen, lagern sich mit Mikrotubuli des andern Poles an
- Metaphase: Es bildet sich eine imaginäre Ebene (Äquatorialplatte) zwischen den beiden Polen, auf der sich die Centromere
der Chromosomen anordnen;
- Anaphase: Die gepaarten Centromere jedes Chromosomes separieren sich; die Tochterchromosomen wandern entlang der Spindelfasern zu den Polen, zur gleichen Zeit entfernen sich
die Pole voneinander, da die nichtkinetochoren Mikrotubuli läger werden
- Telophase: Neue Kernmembran wird gebildet, der Nucleolus erscheint wieder; Spindelapparat wird abgebaut; Chromosomen wandeln
sich in die Arbeitsform um
Die Cytoskinese, also die Teilung des Cytoplasmas, beginnt während den letzten Phasen der Mitose.
Die Kernspindel verteilt die Chromosomen auf die Tochterzellen
Die Kernspindel wird während der Prophase gebildet. Sie besteht aus Mikrotubuli und angegliederten Proteinen.Mehrere parallele Mikrotubuli bilden eine Spindel. Die Ausdehnung geschieht mit Einlagerung
von Tubulin (Protein). Der Aufbau der Mikrotubuli startet in den Centrosomen (Organisationszentrum). Jedes Chromosom besitzt zwei Kinetechore (=Struktur von Proteinen, spezifischer Teil der chromosomalen
DNA am Centromer), die in unterschiedliche Richtungen
blicken. Die Spindelmikrotubuli binden an die Kinetochore und ziehen so die Chromosomen zum Pol. Da die Chromosomen aber von beiden Seiten angezogen werden, entsteht ein Kampf, der schliesslich damit
endet, dass die Chromosomen in der Metaphase auf der Äquatorialplatte zu liegen kommen. Zu Beginn der Anaphase werden die Proteine, welche die Chromatide zusammenhalten, deaktiviert und so können
sie zu den entgegen gesetzten Polen wandern. Die Kinetochore wandern mit Hilfe von Motorproteinen den Miktrotubuli entlang, gleichzeitig werden diese
Mikrotubuli kürzer. Die nichtkinetochoren Mikrotubuli werden länger und dehnen so die Zelle aus.
Cytoskinese
Das erste Zeichen ist eine Teilungsfurche, welche auf der Zelloberfläche in der Nähe der ehemaligen Äquatorialplatte entsteht. Sie bildet sich, da sich ein kontraktiler Ring aus Actinfasern
zusammenzieht. Die Furche wird immer tiefer, bis die Zelle in zwei Teile zerfällt.
Cytoskinese in Pflanzenzellen läuft anders ab. Es gibt keine Teilungsfurche, dafür wird im innern eine Zellplatte aus Zellwandmaterial gebildet, welche das Zellplasma für die zwei Tochterzellen
trennt. Später wird daraus dann eine neue Zellwand gebaut und so werden die Tochterzellen endgültig getrennt.
Mitose in Eukaryoten könnte von der binären Spaltung der Bakterien evolviert haben
Bakterien pflanzen sich fort, indem sie sich einfach teilen. Dies geschieht, indem sich die Chromosomen nach der Teilung sofort voneinander fortbewegen. Wie das geschieht ist jedoch noch nicht bekannt.
Während der Replikation der DNA
vergrössert sich die Zelle bis sie ihre doppelte Grösse erreicht hat. Danach wächst die Plasmamembran in die Zelle hinein und teilt so die Zelle in zwei Tochterzellen.
Da das Ganze bei Eukaryoten mit grösserem Genom und einer Kernmembran nicht mehr so einfach ist, entwickelte sich eine neue Art der Zellteilung. Mögliche Zwischenschritte dieser Evolution hat
man in einzelligen Algen gefunden. Bei
beiden Arten bleibt die Kernmembran intakt. In der einen Art (Dinoflagellaten) sind die replizierten Chromosomen an der Kernmembran befestigt. In der anderen Art (Diatome) separiert eine Spindel im Kern
die Chromosomen.
Regulation des Zellzyklus
Ein molekulares Kontrollsystem steuert den Zellzyklus.
Der Zellzyklus ist durch ein Zellzyklus-Kontrollsystem gesteuert, welches Schlüsselereignisse auslöst und kontrolliert.
Ein wichtiger Teil dieses Kontrollsystems sind die Kontrollpunkte. Kontrollpunkte sind kritische Punkte im Zellzyklus wo Stopp- und Weitersignale den Weitergang des Zyklus bestimmen.
Tierzellen haben eingebaute Stoppsignale, welche den Zyklus stoppen, bis sie von einem Weitersignal aufgehoben werden. Bekannte Kontrollpunkte sind:
- G1 = Restriktionspunkt; liegt in der G1-Phase; hier wird entschieden ob die Zelle geteilt wird oder nicht: wenn hier kein Weitersignal eintrifft, wechselt die Zelle in die G0-Phase.
Das heisst, dass die Zelle den Zellzyklus verlassen hat und nun in einem Status ist in welchem sie sich nicht mehr teilt; die meisten menschlichen Zellen sind in diesem Status, einige Zellen (z.B Leberzellen)
können „zurückgerufen“ werden
- G2 = liegt am Ende der G2-Phase und leitet die M-Phase ein
- M = kontrolliert den Beginn der Anaphase; MPF bricht auseinander
Proteinkinase
Proteinkinasen sind Enzyme welche andere Proteine aktivieren oder inaktivieren indem sie sie phosphorylieren.
Spezielle Proteinkinasen geben das Weitersignal am G1- und am G2- Kontrollpunkt. Diese Kinasen sind in einer wachsenden Zelle immer in einer konstanten Konzentration vorhanden, aber meistens in der inaktiven
Form. Um aktiv zu
sein muss eine solche Kinase an einem Cyclin (= Protein, welches seinen Namen von der zyklischen Schwankung der Konzentration in der Zelle hat) gebunden sein, dadurch sind werden sie cyclin-dependent
kinases genannt (Cdk).
MPF (=maturation-promoting factor) ist der Cyclin-Cdk-Komplex, der auch „M-phase-promoting factor“ genannt wird, da er die Mphase nach dem G2-Kontrollpunkt einleitet. Dies geschieht indem
Proteine phosphoryliert werden und auch der Zerfall der Kernmembran wird durch MPF gesteuert.
Innere und Äussere Signale steuern den Zellzyklus.
Innere Signale: Wenn alle Kinetochore gebunden sind, wird das Stoppsignal beendet. Der Anaphase-Promotor-Komplex (APC) wird aktiviert und verursacht indirekt den Zusammenbruch von Cylcin und die Deaktivierung
von Proteinen, welche die Schwesterchromatide zusammenhalten.
Äussere Signale: Es gibt viele äussere Faktoren (chemische und physikalische), welche die Zellteilung beeinflussen können.
Bsp. 1: Zellen teilen sich nicht, wenn ein essenzieller Nährstoff im Kulturmedium fehlt.
Bsp. 2: die meisten Säugetierzellen teilen sich nur, wenn das Kulturmedium spezifische Wachstumsfaktoren (= Protein, welches von bestimmten Körperzellen entlassen wird und andere Zellen zum
Teilen stimuliert) enthalten.
Wichtig in diesem Zusammenhang ist ein spezieller Wachstumsfaktor, genannt PDGF (plateletderived growth factor), welcher von Blutzellen gemacht wird. Die Bindung von PDGF-Molekülen an Rezeptoren
löst eine Weitergabe von Signalen aus, welche zur Zellteilung führt.
Die Entdeckung von Wachstumsfaktoren führte zum Verständnis der dichteabhängigen Hemmung (= Phänomen, in welchem die beengten Zellen aufhören sich zu teilen). Die Zellen teilen
sich bis sie eine Einzelschicht gebildet haben, dann stoppen sie die Teilung, weil die Menge der Wachstumsfaktoren und der Nährstoffe nicht mehr weiterreicht.
Die meisten Tierzellen weisen auch eine Fixierungsabh舅gigkeit auf. Damit sie sich teilen kömnen, müssen sie an einem Untergrund (z.B Kulturmedium, Äusseres eines Gewebes,...). Die
Fixierung wird dem Zellzyklus-Konrollsystem via Signalweg mitgeteilt, in welchem einbezogene Plasmamembran-Proteine und Elemente des Cytoskeletts eine Rolle spielen.
Krebszellen haben sich von der Kontrolle des Zellzyklus befreit Krebszellen weisen keine dichteabhängige Hemmung oder Fixierungsabhängigkeit auf. Sie stoppen die Zellteilung auch nicht, wenn
keine Nährstoffe mehr vorhanden sind. Wenn sie aber trotzdem aufhören sich zu teilen, machen sie das nicht wie normale Zellen am Restriktionspunkt, sondern einfach zufällig. Krebszellen
sind im Gegensatz zu normalen Zellen dazu fähig, sich unendlich oft zu teilen.
Das Leben einer Krebszelle beginnt mit der Transformation (= Umwandlung einer normalen Zelle in eine Krebszelle). Normalerweise wird eine solche Zelle vom Immunsystem erkannt und zerstört. Wenn
eine Zelle nicht erkannt wurde, bildet sich ein Tumor (= Masse von abnormalen Zellen, in ansonsten normalem Gewebe). Wenn die Zelle am ursprüglichen Ort bleibt, wird die Geschwulst gutartig genannt.
Die meisten gutartigen Tumore verursachen keine Probleme und können komplett beseitigt werden. Dagegen ist ein bösartiger Tumor dazu fähig, ein oder mehrere Organe zu schädigen.
Die Zellen eines bösartigen Tumors können eine unänliche Anzahl an Chromosomen besitzen, ihr Stoffwechsel kann gestört sein und sie verlieren die Bindung an benachbarte Zellen und
können so in anderes Gewebe gelangen. Diese Ausbreitung wird Metastase genannt.

