|
 |
||||||||||||
Informationsvermittlung durch elektrische Erregung
Membranpotentiale können über intrazelluläre Glaskapillarelektroden oder Patch-clamp-Elektroden mit zerstörter Innenmembran gemessen werden. Die Membranpotentiale sind in Ruhe negativ gegenüber dem Extrazellulärraum. Solche Ruhepotentiale liegen bei Nerven- und Muskelzellen von Warmblütern zwischen -55 und -100 mV. Das Ruhepotential ist in erster Näherung ein K+-Diffusionspotential, da in Ruhe vorwiegend die Membrankanäle für K+ offen sind (bei Muskelzellen auch die Cl--Kanäle). Während Zellaktivität kann durch Austritt von K+ aus der Zelle in den schmalen Extrazellulärraum die extrazelluläre K+-Konzentration von normal 3-4 mmol/l auf bis zu 10 mmol/l ansteigen. Dies führt entsprechend der NERNST-Gleichung zur Abnahme des Ruhepotentials (Depolarisation). Starke synaptische Aktivität kann im ZNS die Konzentration an Ca2+ im Extrazellulärraum vermindern und zur Überreregbarkeit führen. Gliazellen sind durch gap junctions miteinander verbunden, so daß Ausgleichströme zwischen depolarisierten und nichtdepolarisierten Gliazellen fließen können. Lokale extrazelluläre Zunahmen der K+-Konzentration (durch erhöhte Neuronenaktivität) werden durch diese Ströme abgepuffert. Die Gliazellen können auch durch Diffusion und Transport Substanzen aus dem Interzellulärspalt aufnehmen. Zusammen mit dem fensterlosen Endothel der Hirnkapillaren bilden sie daher einen wichtigen Bestandteil der Blut-Hirn-Schranke. Die intrazellulären Ionenkonzentrationen werden im wesentlichen von der Na+-K+-Pumpe konstant gehalten, die auch die Metastabilität des Ruhepotentials bedingt.
Aktionspotential
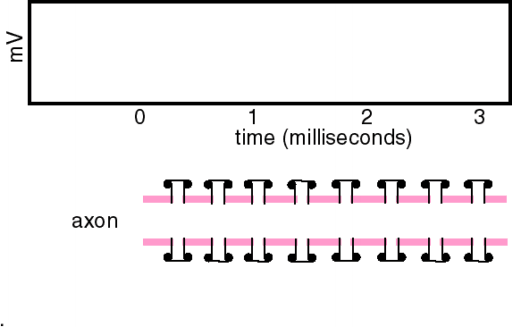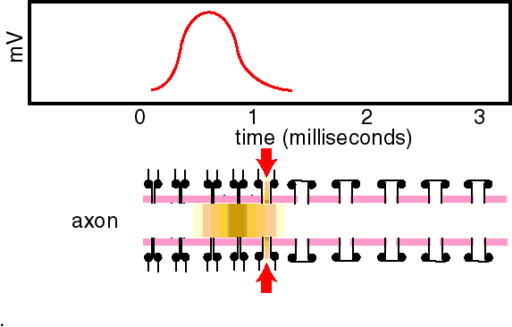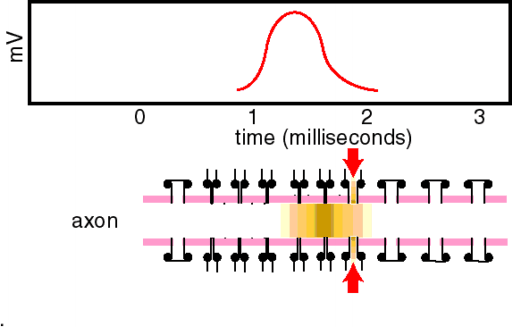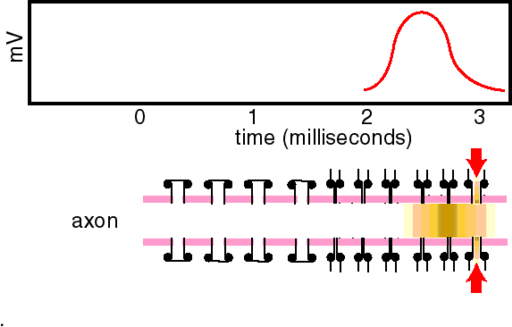
Ein Aktionspotential dauert an Nerven- und Skelettmuskelzellen etwa 1 ms bzw. 10 ms, am Herzmuskel mehr als 200 ms. Es wird ausgelöst, sobald das Ruhepotential zum Schwellenpotential von etwa B50 mV depolarisiert wird und läuft dann stereotyp ab (Alles-oder-Nichts-Prinzip). Der Aufstrich (Depolarisationsphase) dauert dabei nur 0,2-0,5 ms. Die Depolarisation wird durch Na+-Einstrom (als Folge einer vorübergehend erhöhten Na-Leitfähigkeit) verursacht, die Repolarisation durch K+-Ausstrom beschleunigt (durch eine ebenfalls , wenn auch leicht verzögerte Erhöhung der K-Leitfähigkeit). Die Analyse der Leitfähigkeitsänderungen während des Aktionspotentials ist mit Hilfe der Spannungsklemme (voltage clamp) möglich. Zur Analyse der Na+-Ströme werden die K-kanäle mit TEA blockiert, zur Analyse der K+-Ströme die Na-Kanäle mit TTX. Aus den Meßergebnissen können die Zeitverläufe der Membranleitfähigkeiten g bestimmt werden. Nach Depolarisation zur Schwelle steigt gNa sofort und schnell aber nur kurzzeitig an (Inaktivation). Ebenfalls depolarisationsbedingt, aber verzögert, und ohne Inaktivation, erhöht sich auch gK. Auch gCa steigt bei Depolarisation. Nach dem Aufstrich des Aktionspotentials inaktiviert der Na-Strom. Für etwa eine Millisekunde nach dem Aktionspotential ist die Membran deshalb auch durch große Depolarisationen nicht erregbar, sie ist absolut refraktär. Anschließend, mit zurückgehender Inaktivierung, ist die Membran einige Millisekunden vermindert erregbar, also nur noch relativ refraktär. Durch die Refraktärität ist die maximale Entladungsfrequenz von Neuronen auf 500 bis 1000 Aktionpotentiale pro Sekunde begrenzt. Aktionspotentiale sind häufig von Nachpotentialen gefolgt. Hyperpolarisierende Nachpotentiale sind teils durch eine etwas überdauernde gK, teils durch vermehrtes Auspumpen von Na+ verursacht. Bei Erhöhung der Ca2+-Außenkonzentration verschiebt sich das Schwellenpotential zu positiveren Werten, die Zelle wird weniger leicht erregbar. Erniedrigung der Ca2+-Außenkonzentration hat die umgekehrten Effekte (Gefahr von epileptiformen Neuronenentladungen und Muskelkrämpfen). Die „patch clamp“ ermöglicht die Messung von Kanalströmen durch einzelne Kanalmoleküle. Die Stromamplitude liegt im pA-Bereich, die -dauer um 1 ms. Nach Depolarisation öffnen Na+-Kanäle einmal, und zwar und der ersten Milisekunde. die Öffnung erfolgt nicht streng determiniert, sondern stochastisch. Nach der Öffnung geht der Kanal in einen geschlossen-inktivierbaren Zustand über, bevor er wieder in den geschlossen-inaktivierbaren Zustand zurückkehrt (entspricht der Inaktivation des Na-Stroms bzw. der Refraktärität). Die die Repolarisation des Aktionspotentials bewirkenden K+-Kanäle öffnen wiederholt während der ganzen Depolarisationsdauer. Es gibt auch andere, z.B. inaktivierende K+-Kanaltypen. Ca2+-Kanäle öffnen nach Depolarisation in kurzen Salven (bursts), aber relativ selten. Ihre Öffnungswahrscheinlichkeit wird durch cAMP in der Zelle heraufgesetzt (Modulation, z.B. an Herzmuskelzellen bewirkt durch Adrenalin). Mit molekularbiologischen Methoden können Kanalcharakteristika, wie Ionenselektivität, potentialabhängige Öffnung und Inaktivation, bestimmten Molekülanteilen zugeordnet werden. Dies geschieht z.B. durch den Austausch von Aminosäuren durch gezielte Mutationen oder Blockade von Aminosäuresequenzen durch Antikörper. Alle Kanalproteine stammen möglicherweise vom Ca-Kanalmolekül ab. Am besten bekannt ist der Na-Kanal, ein großes Glykoprotein mit einem Durchmesser von 8 nm und einer Kanalöffnung von etwa 0,5 nm. Seine Blockade durch Lokalanästhetika unterbricht die Fortleitung von Aktionspotentialen (z.B. lokaler Nervenblock zur Schmerzausschaltung bei zahnärztlichen und chirurgischen Eingriffen). Werden in einer Zelle Stromstöße appliziert, so lösen diese elektrotonische Potentiale aus. Bei homogener Stromverteilung in kugeligen Zellen steigen die elektrotonischen Potentiale negativ-exponentiell [?] mit der Membranzeitkonstante t an. Bei langgestreckten Zellen nimmt die Amplitude des elektrotonischen Potentials negativ-exponentiell mit der Entfernung mit der Membranlänskonstante l ab. extrazelluläre, z.B. auf die Haut über einem Nerven applizierte Stromstöße depolarisieren die Axone nahe der Kathode und hyperpolarisieren nahe der Anode. Wenn die Depolarisationen überschwellig werden, lösen die an der Kathode Aktionspotentiale aus. Von außerhalb der Zelle eingespeiste, überschwellige, depolarisierende Ströme werden Reize genannt. Hochfrequente Ströme fließen nicht lange genug in eine Richtung, um überschwellig zu depolarisieren; sie erwärmen lediglich das Gewebe (wird bei der Diathermie ausgenutzt). Aktionspotentiale werden mit unverminderter Amplitude in Nervenfasern fortgeleitet (Alles-oder-Nichts-Prinzip). Die Leitungsgeschwindigkeit ist bis zu 100 m/s in dicken markhaltigen Axonen, und kleiner 1 m/s in dünnen marklosen Fasern. Das Aktionspotential, das eine Stelle des Axons erreicht hat, depolarisiert elektrotonisch den vor ihm liegenden Axonabschnitt. Wenn diese Depolarisation die Schwelle erreicht, wird wiederum ein Aktionspotential ausgelöst, und der Erregungsvorgang hat sich damit vorwärts bewegt. Markhaltige Fasern sind praktisch nur an den Schnürringen erregbar; über die Internodien breitet sich die Depolarisation elektrotonisch und fast ohne Zeitverlust aus, weil die Myelinscheide den effektiven Membranwiderstand erhöht und die Kapazität herabsetzt (saltatorische Erregungsleitung). Damit wird die Fortleitung in markhaltigen Fasern relativ zu den marklosen beschleunigt. Bei markhaltigen wie marklosen Fasern steigt die Leitungsgeschwindigkeit mit der Zunahme des Faserdurchmessers. An Sensorzellen der Sinnesorgane oder an Synapsen werden längerdauernde Depolarisationen erzeugt, die die Amplitude von Signalen abbilden. Erregbare Zellmembranen können solche Depolarisationen in Serien von Aktionspotentialen umsetzen, die in Axonen zur nächsten Synapse weitergeleitet werden. Auf diese Umsetzung spezialisierte Membranbezirke enthalten verschiedene Typen von K-Kanälen, die die Repolarisation der Aktionspotentiale so modifizieren, daß Impulsserien verschiedener Frequenz und Dauer, die der Größe der Dauerdepolarisation entsprechen, ausgelöst werden. |
|||||||||||||
| Seitenanfang | Kontakt | Newsletter | Impressum | |||||||||||||