|
 |
||||||||||||
Zellphysiologie
Jede Zelle ist von einer Membran umschlossen, die aus Lipiddoppelschichten aufgebaut ist. Diese werden von Phosphatidylcholinmolekülen gebildet. In die Doppelschicht sind weitere Lipide, nämlich Glykolipide (an der Außenseite) und Cholesterin (zur Membranstabilisierung) eingelagert und v.a. Proteine, die entweder nur an der äußeren oder nur an der inneren Schicht verankert sind oder sich durch beide Schichten erstrecken. Die in der Membran liegenden Teile sind hydrophob, die äußeren hydrophil. In der Zelle befinden sich zu gleichen Volumenteilen membranumschlossene Organellen (v.a. endoplasmatisches Retikulum, Zellkern, Mitochondrien) und das Zytosol, das v.a. gelöste Eiweiße und kleine Ionen enthält. Das Zytosol wird vom Zytoskelett durchzogen, das aus Mikrotubuli, Aktinfilamenten und intermediären Filamenten aufgebaut ist. In der Zelle ist die Na+-Konzentration mehr als 10mal geringer als in der Außenlösung; die K+-Konzentrationen verhalten sich umgekehrt. Die intrazelluläre Ca2+-Konzentration beträgt etwa 0,1 µmol/l. In Flüssigkeiten wie dem Extrazellulärraum und dem Zytosol gleichen sich Konzentrationsunterschiede von gelösten Stoffen durch Diffusion aus (1. Fick-Diffusionsgesetz). Auch die Lipidmembran kann von einer Reihe von Molekülen permeiert werden, v.a. von Wasser, O2, CO2 und lipidlöslichen Stoffen. Für die meisten Moleküle, insbesondere alle geladenen, ist die Lipidmembran aber impermeabel.
Die K+-, Na+-, ClB- und Ca2+-Ionen diffundieren daher durch wassergefüllte Poren in Kanalproteinen der Membran, getrieben durch ihren Konzentrationsgradienten und die Differenz zwischen dem Membranpotential (Ruhepotential) und ihrem spezifischen Gleichgewichtspotential.
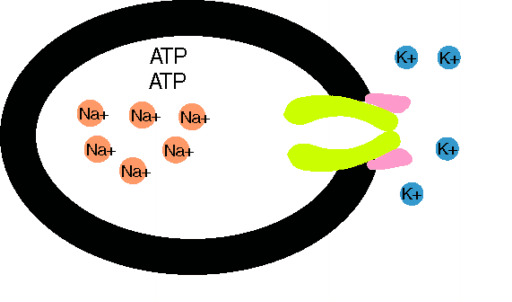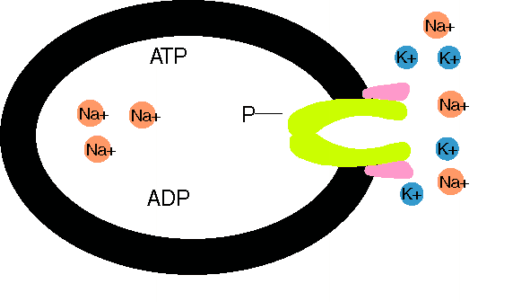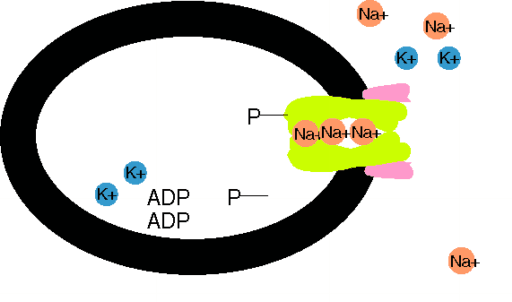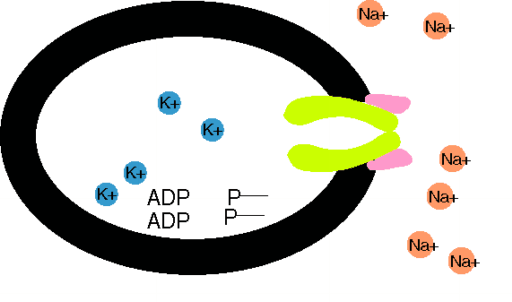
Das Ungleichgewicht der Ionenflüsse wird durch aktiven Transport kompensiert. Der wichtigste ist die Na+-K+-Pumpe, eine ATPase an der Innenseite der Membran, die für den Verbrauch von 1 Molekül ATP (Spaltung in ADP + Phosphat) 3 Na+ aus der Zelle und 2 K+ in die Zelle schafft.Für diese elektrogene Pumpe wird 1/3 des Zellenergieverbrauchs aufgewendet. Die Membranpumpen sorgen für die Metastabilität des Ruhepotentials und die Konstanz des Zellvolumens. Der durch die Pumpaktivität aufgebaute Na+-Konzentrationsgradient über der Zellmembran treibt sekundär aktive Transporte, z.B. von Ca2+, Zuckern und Aminosäuren, in die (Symporte) und aus der Zellen (Antiporte). Makromoleküle, die über keine Kanaltransportsysteme verfügen, aber z.B. auch Cholesterin, können die Zellmembran durch Endo- oder Exozytose kreuzen.
Na+-K+-Pumpe
Im Zytosol und in den von den Organellen eingeschlossenen Flüssigkeitsräumen werden Konzentrationsgradienten durch Diffusion ausgeglichen. Die Organellenmembranen verfügen darüberhinaus über die gleichen aktiven Transportmechanismen wie die Zellmembran. Verschiedenste Stoffe, z.B. neu synthetisierte Proteine oder Cholesterin, werden intrazellulär in Vesikeln kompartimentiert, transportiert und über die Zell- und Organellenmembranen geschleust. Gerichtete Transporte durch Auf- und Abbau von Organellen und durch Umlagerungsprozesse verändern die Zellform, nehmen am Wachstum teil und bewegen Vesikel und Organellen entlang von Mikrotubuli und Filamenten. Der axonale Transport hat verschiedene Komponenten, unter ihnen den schnellen
anterograden Transport vom Zellkörper zu den Nervenendigungen mit einer
Geschwindigkeit von 410 mm/Tag, wie auch langsamere anterograde und retrograde
Transportmechanismen. Zellen sind keineswegs die statischen Gebilde, als die
sie notwendig z.B. im elektronenmikroskopischen Schnitt erscheinen. die Plasmamembran
und erst recht die Organellen sind ständig in rascher Bewegung und Umbau
und erfüllen erst so ihre Funktionen. sie sind auch nicht schlichte Reagenzgefäße
für chemische Reaktionen, sondern hochorganisierte Konglomerate von Membranen
und Fibrillen, an denen Reaktionen in optimal geordneter Form ablaufen können. |
|||||||||||||
| Seitenanfang | Kontakt | Newsletter | Impressum | |||||||||||||
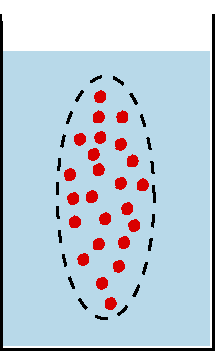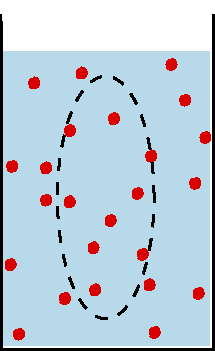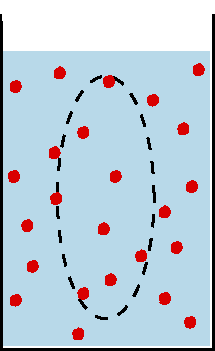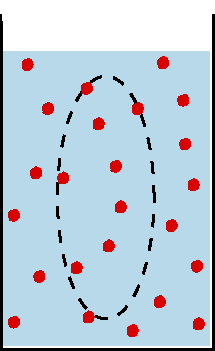 Beim Ruhepotential liegt das Potential nahe dem K+-Gleichgewichtspotential, also demjenigen Potential,
bei dem die Wirkung des durch die Auswärtsdiffusion von K+-Ionen aufgebauten
elektrischen Feldes den von der Konzentrationsdifferenz aufgebauten Diffusionsdruck
kompensiert (Nernst-Gleichung). die überwiegende Rolle der K+-Ionen für
das Ruhepotential ist darauf zurückzuführen, daß in der ruhenden
Membran geöffnete K+-Kanäle weitaus häufiger sind als solche
für Na+ oder ClB.
Beim Ruhepotential liegt das Potential nahe dem K+-Gleichgewichtspotential, also demjenigen Potential,
bei dem die Wirkung des durch die Auswärtsdiffusion von K+-Ionen aufgebauten
elektrischen Feldes den von der Konzentrationsdifferenz aufgebauten Diffusionsdruck
kompensiert (Nernst-Gleichung). die überwiegende Rolle der K+-Ionen für
das Ruhepotential ist darauf zurückzuführen, daß in der ruhenden
Membran geöffnete K+-Kanäle weitaus häufiger sind als solche
für Na+ oder ClB.